





Abstract
Chitotriosidase (CHIT1) is a highly conserved and regulated chitinase secreted by activated macrophages; it is a member of the 18-glycosylase family (GH18). CHIT1 is the most prominent chitinase in humans, can cleave chitin and participates in the body's immune response and is associated with inflammation, infection, tissue damage and remodelling processes. Recently, CHIT1 has been reported to be involved in the molecular pathogenesis of pulmonary fibrosis, bronchial asthma, COPD and pulmonary infections, shedding new light on the role of these proteins in lung pathophysiology. The potential roles of CHIT1 in lung diseases are reviewed in this article.
Abstract
This is the first review of chitotriosidase in lung disease http://bit.ly/2LpZUQI
Introduction
Chitinases are glycosylated hydrolytic enzymes that catalyse the degradation of chitin into N-acetylglucosamine, which is an important carbon and nitrogen source. These enzymes are expressed widely in plants, bacteria, fungi, actinomycetes, insects and animals including humans [1]. Chitin was first reported by Benecke [2] in 1905, who isolated Bacillus chitinivorous which dissolves chitin and utilises it as a nutrient (figure 1). 16 years later, Folpmers [3] observed hydrolytic circles in plates containing chitin after adding two strains of microorganisms isolated from the harbour water at Kiel, which further proved the existence of chitinases. Finally, in 1929, Karrer and Hofmann [4] extracted the enzyme from the land snail Helix pomatia. Based on the homology of amino acids in chitinases, they were divided into multiple gene families including 18-, 19- and 20-glycosylase families [1, 5, 6]. Subsequently, glycoside hydrolase families 23, 45 and 48 were reported [7–9]. Both families 18 and 19 comprise chitinases from viruses, bacteria, fungi, animals and certain plants. However, these families have been said to evolve from different ancestors due to the lack of amino acid sequence homology and three-dimensional structure among these chitinases. Family 20 glycosylase enzymes include the N-acetylglucosaminidases from bacteria and certain fungi [10].

Enzymatic activity of chitotriosidase (CHIT1) in organisms.
Chitinases play various roles in different organisms. Most bacteria produce chitinases to degrade chitin in order to supply carbon and nitrogen as an energy source for their growth and replication [3, 11, 12]. In plants they play a protective role against foreign pathogens such as fungi and insects [13]. Chitinases of insects are mainly related to their late embryonic development and ecdysis [14]. Although mammals, including humans, do not have any endogenous chitin, they express chitinases and chitinase-like proteins, which are members of the evolutionary conserved family of 18 glycosyl hydrolases with high homology in structure, and can be further divided into active chitinases (chitotriosidase (CHIT1) and acidic mammalian chitinase (AMCase)) and inactive chitinase or chitinase-like proteins (Chi3L1/YKL40, Chi3L2/YKL39, Chi3L3/YM1, Chi3L4/YM2, Chi3L5/YM3, Chi3L6/YM4, Chi3L7/Bclp1/Bclp2, SI-CLP, OVGP1) [15–18] (table 1). While both chitinases and chitinase-like proteins can bind to chitin with high affinity, only true chitinases can cleave it.
Representative members of the conserved family of 18 glycosyl hydrolases
Mammals have two true chitinases: CHIT1 and AMCase. CHIT1 is the most prominent chitinase in human lung and circulation, where all the chitinase activity is attributed to CHIT1, while the contribution of AMCase in humans is negligible [19–22]. CHIT1 gene is located in chromosome 1q32.1, consisting of 13 exons and spanning ∼20 kb to encode a 445 amino acid long protein [23, 24]. CHIT1 is believed to be secreted by activated macrophages, although even at baseline there is abundance of chitinase proteins in the lungs and intestine, suggesting some baseline secretion by resident macrophages or structural cells or both [25, 26]. CHIT1 has been shown to participate in the innate and acquired immune responses [18, 27–30], and have been reported to correlate with various diseases including cancer, diabetes, asthma, sarcoidosis, infections, inflammatory bowel disease, atherosclerosis and nervous system diseases [26, 31–36].
Biological roles of CHIT1
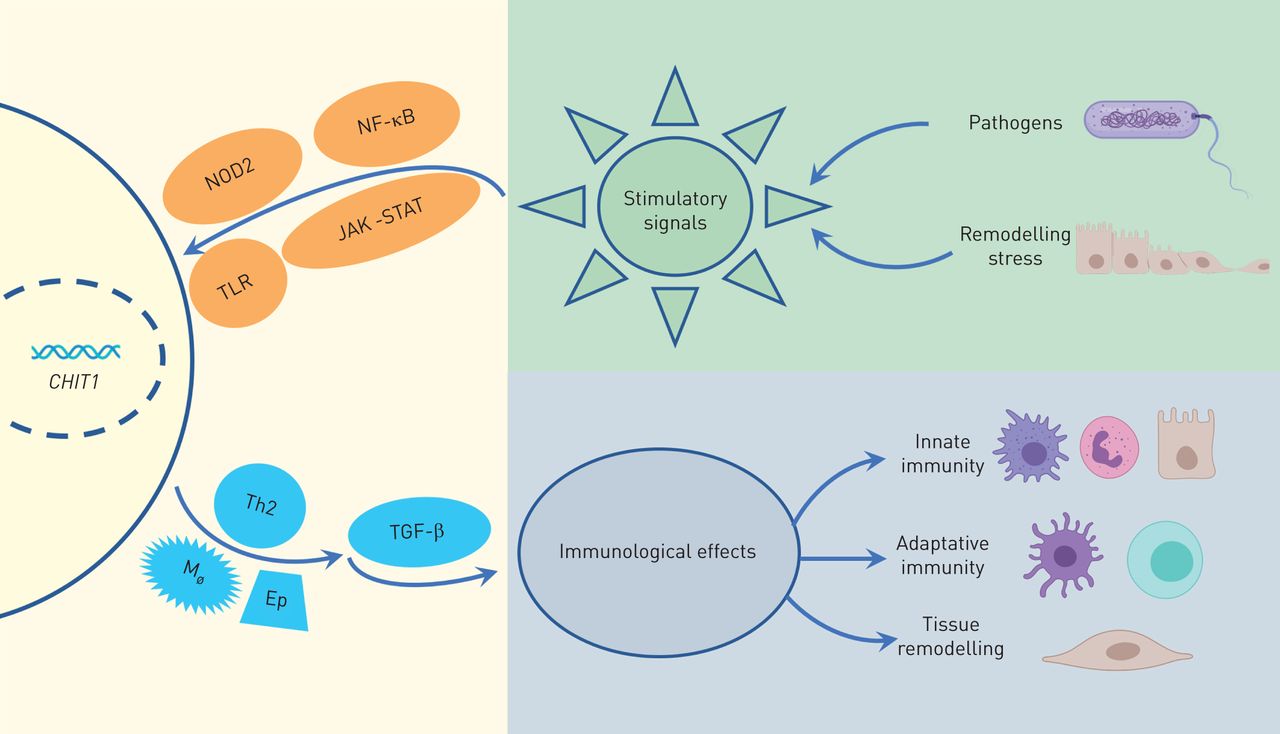
CHIT1 was the first chitinase to be discovered and characterised in mammals [23, 37]. Both immune cells, such as activated macrophages and neutrophils, and structural cells, such as epithelial cells can secrete CHIT1 [26, 37–41]. These cells constantly face environmental threats given their location in the lung, suggesting its role in innate immune system in pulmonary diseases [26, 37, 40, 42]. Macrophages and neutrophils release CHIT1 after stimulation with interferons, tumour necrosis factor (TNF) and granulocyte-macrophage colony stimulating factor through NF-κB, Jak-Stat, NOD2 and Toll-like receptor signalling pathways [38, 43–46], indicating the importance of this enzyme in immune response [38, 46]. Furthermore, CHIT1 is stored in ready-to-release granules in the immune cells along with other antimicrobial agents such as lysozyme, again emphasising the possible roles of CHIT1 in immunity [47] (figure 2).

Biological activity of chitotriosidase (CHIT1). TLR: Toll-like receptor; Th2: T-helper type 2 cell; TGF: transforming growth factor.
The basic biological function of chitinase is that it can hydrolyse chitin. Based on this property, it is believed to play an important role in biological defence against or digestion of pathogens containing chitin as part of their structure, such as fungus and arthropods. However, due to the evolutionary conserved nature of this protein, a wide variety of functions has been demonstrated across species for chitinases and related proteins [48]. For example, the CHIT1-homologous genes, such as imaginal disc growth factor (IDGF) from Drosophila, My-Clp1 from Japanese scallops and Cg-Clp1 and Cg-Clp2 from oysters have been shown to be in involved in wound healing, developmental tissue remodelling and immune defence [49–53]. In Drosophila, IDGF is essential for larval and adult moulting as it organises extracellular matrix formation at barrier tissues [54]. In Japanese scallops, the expression of My-Clp1 was observed in various developmental stages of metamorphosis, which suggest its roles in tissue degeneration and remodelling [55]. In oysters, Cg-Clp1 and Cg-Clp2 were reported to be involved in immune responses to bacterial lipopolysaccharide challenge [53].
However, the precise function of CHIT1 in mammals still remains unknown. CHIT1 does not appear to play a role during embryogenesis or early development in mammals as CHIT1-deficient mice have been developed successfully and do not show any overt signs of abnormalities [36]. Most of the clinical studies looking at the role of CHIT1 have largely been correlational. Basic science studies have shown several mechanistic insights into the role of CHIT1 during various disease models. In this review, we focus on the lung diseases and possible roles of CHIT1 (figure 3).

Chitotriosidase (CHIT1) in respiratory disorders. IL: interleukin; TGF: transforming growth factor.
CHIT1 and interstitial lung diseases
Interstitial lung diseases (ILDs) are a group of disorders with basic pathological changes including diffused parenchymal injury, alveolitis and fibrosis, with clinical manifestations such as shortness of breath, diffused shadow shown on chest radiography, restrictive ventilatory impairment, reduced diffusing capacity of the lung for carbon monoxide and hypoxaemia [56, 57]. The aetiology and pathogenesis of various ILDs are still not entirely known. The accumulation of collagen, secreted by myofibroblasts that are recruited in response to hypothesised microinjuries of the alveolar epithelium, leads to the development and sustainment of the fibrotic process [58]. Chitin is one of the abundantly present substances in the environment that can be inhaled into the lung and directly damage the alveolar epithelial cells [59, 60]. Chitins such as those present in the house dust mite can easily reach the lungs by inhalation. Van Dyken et al. [19] found that the amounts of chitin polymer content are increased in bronchoalveolar (BAL) fluid from patients with ILD compared to healthy controls, suggesting a possible role of impaired chitin clearance in the pathogenesis of pulmonary interstitial fibrosis. In other studies, CHIT1 concentrations in BAL were significantly increased in patients with sarcoidosis and idiopathic pulmonary fibrosis (IPF) compared to healthy BAL, although a consistent increase was only observed in the serum of patients with sarcoidosis, indicating compartment-specific regulation of CHIT1 and its possible role in causing pulmonary tissue remodelling and tissue damage in interstitial diseases [61–63]. However, studies have not yet reported whether there is a clear link between the amount of chitin in the lung and levels of chitinase, and if any of these two or both can possibly explain disease onset, progression and severity. In addition, recent studies have demonstrated increased CHIT1 expression in single-cell transcriptomes of an IPF-specific macrophage subpopulation [64, 65].
Mechanistic studies have shed important light on the role of CHIT1 in various cellular signalling pathways that can explain their contribution to ILDs. During the fibrotic process, CHIT1 enhanced transforming growth factor (TGF)-β-stimulated fibroblast proliferation and myofibroblast transformation through interaction with Tgfbrap1 and FoxO3a [66]. This led to the enhancement of TGF-β-stimulated Smad2/3 and MAPK/Erk activation and inhibition of TGF-β-stimulated Smad7 expression, which acts as a negative regulator of TGF-β signalling. At the same time, the expression of CHIT1 was increased in the lungs or blood of patients with IPF and scleroderma-associated ILD patients where CHIT1 expression was inversely associated with Smad7 expression [67]. Furthermore, bleomycin-induced pulmonary fibrosis was significantly reduced in CHIT1−/− mice, which may be due to CHIT1-mediated activation of fibroblasts by augmenting TGF-β1 and TGF-β1 receptors which induced the Smad and MAPK/ERK signalling pathways [40]. Therefore, it is conceivable that CHIT1 plays a role in the pathogenesis of IPF. A study by Dymek et al. [68] showed that CHIT1 was highly expressed in the macrophages of IPF patients. Based on this evidence, therapeutic inhibition of CHIT1 has been explored to alleviate lung fibrosis. In the bleomycin-induced pulmonary fibrosis model, the CHIT1 inhibitor OATD-01 had more significant therapeutic effects compared to treatment with nintedanib (an antifibrotic agent), while the therapeutic efficiency was comparable to pirfenidone. Both nintedanib and pirfenidone are currently approved for treatment of patients with IPF [68, 69]. If successful, these therapies will put chitinase in a strong spotlight as a modulator of human diseases.
CHIT1 and pulmonary inflammatory diseases
Pulmonary inflammatory diseases involve multiple inflammatory cells and are manifested as structural changes in the airway wall and/or parenchyma, including asthma, COPD, bronchopulmonary dysplasia and cystic fibrosis [70]. Studies have shown that chitinase plays an important role in airway inflammatory diseases by regulating the inflammatory response through degradation of chitin or pathways independent of chitin degradation [15, 70, 71]. COPD is caused by an abnormal inflammatory response to the inhalation of harmful substances, such as smoking. The upregulation of the CHIT1 gene, especially in macrophages, results in the higher levels of chitinase activity in smokers, which may be involved in COPD pathogenesis and progression of the disease [20]. In bronchial biopsies of smokers with COPD, more chitinase-positive cells and increased proportions of alveolar macrophages expressing CHIT1 were observed [72]. The expression level of chitinase in BAL fluid of patients with COPD were associated with airflow obstruction, emphysema, interleukin (IL)-1β, IL-8, TNF-α and its type II soluble receptor TNF-RII [72]. Thus, increased CHIT1 levels seem to be a risk factor for COPD. Agapov et al. [73] found that the level of plasma CHIT1 were associated with the degree of decline in lung function and increased significantly in severe and very severe COPD, based on which the CHIT1 was thought to be a marker for quantitative stratification of COPD. Genetic association studies suggested that the different CHIT1 genotypes were related with the rate of decline of the baseline forced expiratory volume in 1 s [74].
Similarly, CHIT1 has been implicated in the pathogenesis of asthma; however, the data have been largely conflicting. James et al. [75] reported that CHIT1 levels are significantly elevated during asthma. Furthermore, the level of CHIT1 correlated with disease severity [75]. In contrast, Seibold et al. [20] found that chitinase activity itself in the lung was mostly contributed by CHIT1 and was lower in subjects with asthma compared to the healthy controls. These differences may be due to different exposure profiles in the subjects. Similar to chitinase levels in asthma, conflicting results of genetic variants of CHIT1 and severity of the disease have been reported. In a small study, six cases of children with severe asthma had decreased chitinase expression and activity and all six patients were heterozygous for a 24-bp duplication in the CHIT1 gene [32]. However, Wu et al. [76] found that single nucleotide polymorphisms of the CHIT1 gene were not associated with asthma, asthma-related pulmonary physiological changes or allergen-related phenotypes, according to data from the Childhood Asthma Management Program. In spite of the conflicting evidence regarding the role of CHIT1 in the pathogenesis of airway inflammatory diseases, OAT-889, the inhibitor of CHIT1, has been tested in a mouse model of asthma [77]. These studies demonstrated that OAT-889 had profound anti-inflammatory effects in a chronic asthma model in mice [77]. Mechanistically, CHIT1 was reported to play a protective role in asthma by regulating expression of TGF-β and Foxp3+ regulatory T cells (Tregs). Decreased expression of TGF-β and a decreased number of Foxp3+ Tregs were observed in CHIT1-knockout mice in an asthma model [78]. TGF-β1 stimulation of Treg accumulation is regulated by the ability of CHIT1 to enhance the ability of TGF-β1 to stimulate T-cell TGF-β1 receptor expression. Based on these results, the authors concluded that CHIT1 decreased pulmonary inflammation through modulating T-helper type 2 cell (Th2) responses [78]. Some studies reported that AMCase rather than CHIT1 was involved in the pathogenesis of bronchial asthma in Caucasian children [79, 80]; however, given the fact that most chitinase activity in humans is contributed by CHIT1, the significance of AMCase in modulating diseases remains uncertain.
The significance of CHIT1 in patients with cystic fibrosis does not seem to be evident; however, some studies have indicated a possible relationship between chitinase and cystic fibrosis. Livnat et al. [81] investigated the relationship between CHIT1 duplication in cystic fibrosis patients and their predisposition to allergic bronchial pulmonary aspergillosis. They found there was no strong correlation between CHIT1 duplication (loss of chitinase activity) and W1282X cystic fibrosis transmembrane conductance regulator CFTR mutation [81]. Work by Hector et al. [82] demonstrated that CHIT1 levels were elevated in patients with cystic fibrosis, especially in patients with fungal infections. Additionally, this study demonstrated the susceptibility of CHIT1 to neutrophil elastase mediated cleavage. Given the persistent neutrophilic inflammation in cystic fibrosis lungs, it is not clear how this cleavage of CHIT1 alters its biological activity or its possible contribution to disease progression.
CHIT1 and pulmonary infections
Pulmonary infection manifests as inflammation in the lung caused by pathogenic microorganisms such as bacteria, viruses, fungi or parasites [83, 84]. Pulmonary infection, which affects ∼450 million people worldwide and kills ∼4 million people every year, is one of the leading causes of human deaths and of the ultimate cause of death for people with other serious chronic diseases [85, 86]. As the current population is ageing, infections pose a serious threat to humankind. Ageing leads to immunosuppression, and given the rise of drug-resistant strains of pathogens, the therapeutic options remain limited [87–90]. The role of CHIT1 in infection has been studied [21, 36, 91–93]. Anecdotal evidence has suggested important roles for CHIT1 in host immunity, given its high concentration in anatomical sites such as the lungs and gut where there is constant exposure to microorganisms [19, 94]. Furthermore, CHIT1 is released by activated macrophages and neutrophils, two prominent immune cells that contribute extensively to antiviral and antibacterial immunity [27, 95]. However, experimental evidence for their direct roles in pulmonary infections has only recently started to emerge.
Pulmonary tuberculosis, caused by Mycobacterium tuberculosis, is an infectious disease that attacks the lung [96, 97]. It has been shown that CHIT1 expression levels are increased and correlate with the severity of the disease. CHIT1 is released into the serum as a result of the pulmonary granuloma and its level is reduced with the regression of the inflammation, which may reflect the stage, severity and response to treatment of tuberculosis [63, 98, 99]. Similarly, elevated chitinase activity was observed in patients with cystic fibrosis, especially upon colonisation with Candida albicans. In contrast to chronic infections, acute infections such as Klebsiella pneumoniae or Pseudomonas aeruginosa led to the precipitous decrease of chitinase activity in the BAL. This effect was mediated by neutrophil elastase mediated cleavage of CHIT1 [36]. Similarly, in patients with cystic fibrosis it was observed that neutrophil elastase can cleave CHIT1 protein in a time-dependent manner [82].
As the breakdown of chitin is an obvious function that can work as a host defence mechanism against fungal infections, Gordon-Thomson et al. [100] studied the antifungal properties of chitinase in vitro by transfecting Chinese hamster ovary cells with the CHIT1 gene. The growth of Aspergillus niger, C. albicans and Cryptococcus neoformans can be inhibited by the supernatant of transfected cells. In the mouse model, this study demonstrated that the engraftment of microcapsules containing cells transfected with the chitinase gene have the potential to fight against infection caused by chitin-containing microbes by extending the delivery time of recombinant chitinase. Supporting these experimental models, Hector et al. [82] found that patients with CHIT1 deficiency due to genetic mutation had higher colonisation with C. albicans. However, this study found no association between chitinase genetic deficiency and colonisation of Aspergillus fumigatus, another chitin-containing fungal pathogen. Using a mouse model, Wiesner et al. [93] showed that CD11b+ conventional dendritic cells present chitin recognition via CHIT1 to induce Th2 differentiation during pulmonary cryptococcal infection, which led to pathogenic inflammation. CHIT1-deficient mice had improved survival in the face of cryptococcal infection in this study. These studies show the complexity of the regulation and role of chitinase during pulmonary fungal infections.
There are few studies on the relationship between CHIT1 and pulmonary infection with bacterial infections. We studied the role of CHIT1 during K. pneumoniae infection [36]. CHIT1-/- mice had decreased bacterial dissemination and increased survival of mice during pulmonary Klebsiella infection in the presence or absence of antibiotics [36]. The beneficial effect of CHIT1 deficiency on the host was associated with Akt pathway activation. Thus, targeting CHIT1 may be a new potential therapeutic target for defence against K. pneumoniae. However, these effects seem to be pathogen-specific, as CHIT1-/- mice were more susceptible to Pseudomonas lung infections. These studies indicate that role of CHIT1 is dependent on the type of bacterial infection.
CHIT1 and other lung diseases
Besides the aforementioned diseases, the role of CHIT1 has been indicated in many other lung diseases. Gołab et al. [101] reported that CHIT1 activity in lung cancer patients was higher compared to those with normal controls. However, similar elevations were found in patients with pulmonary exudates without cancer. They concluded that CHIT1 activity in patients' serum and exudate cannot be used as a parameter to distinguish lung cancer from inflammation [35, 101]. Obstructive sleep apnoea (OSA) is a sleep disorder caused by complete or partial obstruction of the upper airway and is characterised by recurrent nocturnal breathing cessation [102]. It has been reported that the prevalence of 24-bp duplication of the CHIT1 gene was higher in a severe sleep apnoea group than controls, and the concentration of CHIT1 was related to age, creatine and testosterone [103]. In a study by Teitsdottir et al. [104], plasma CHIT1 activity was measured before and after treatment with positive airway pressure in 97 OSA patients with an apnoea–hypopnoea index ≥15 events·h−1. The association between OSA severity and CHIT1 activity has not been confirmed. However, the level of CHI3L1 was decreased in severely obese OSA patients after positive airway pressure treatment, but not in mildly obese patients.
Evidence of limited roles of chitinases
CHIT1 is present at high concentrations in mammals at locations that are highly susceptible to invading pathogens, such as foodborne and airborne pathogens infecting the gut and lungs, respectively. Their presence in immune cells in these organs led to the hypothesis that they may have antibacterial properties. Hall et al. [105] explored the role of CHIT1 in killing a wide range of pathogens, either alone or in combination with lysozyme. Contrary to conventional belief, CHIT1 had no direct antibacterial properties against Gram-positive or Gram-negative pathogens, alone or in combination with lysozyme.
The most striking elevation of CHIT1 is observed in lysosomal storage diseases such as Gaucher's disease, where it serves as a strong diagnostic tool for monitoring disease severity and therapeutic effectiveness [42]. Gaucher's disease manifests as multiorgan disease where lung involvement is rare, but present in a subset of patients [106, 107]. However, not many systemic studies have explored directly the specific effects of CHIT1 on various organ phenotypes including in the lungs in these patients. The current knowledge implicating CHIT1 on a specific phenotype in Gaucher's disease is not adequate and requires further direct studies to determine how elevated levels of CHIT1 contribute to lung disease pathogenesis [108–111]. It is still possible that CHIT1 contributes to other organ effects in Gaucher's disease. One possibility is that it may contribute to liver fibrosis, as seen in some of the patients with the disease [112]. Contribution to fibrosis can be explained biologically given the ability of CHIT1 to modulate the TGF-β pathway; however, there are no direct studies to implicate CHIT1 in this phenomenon observed in Gaucher's disease. These observations are a strong reminder of the possible limitations of reverse genetics to study the impact of genes using mouse models where an alteration in one gene can change a cascade of effects, obscuring the effects of the gene per se in an unperturbed natural environment.
Furthermore, upregulation of CHIT1 in many diseases has been controversial and data have been shown to be conflicting. As mentioned earlier, studies have shown both upregulation and downregulation of CHIT1 activity during asthma [108, 113–115]. During fungal infection, upregulation was observed with Candida, but not with Aspergillus, although both fungal agents contain chitin as an integral part of their structure [116, 117]. The most accepted upregulation has been observed only in Gaucher's disease, where its role in disease pathogenesis or protection has not been identified.
Further major evidence suggesting the limited role of biological activity of CHIT1 is that a significant proportion of the human population is deficient in chitinase activity due to a 24-bp mutation in exon 10. The deficiency ranges from 5% to as high as 30% in some populations [110, 118, 119]. In spite of the widespread presence of this deficiency, no overt effect of CHIT1 deficiency has been noted in any epidemiological studies. Studies showing evidence of its role in diseases such as filarial disease are performed in a limited number of subjects and have not been validated extensively [120, 121]. This is surprising given the fact that CHIT1 is the only true chitinase present in the humans, and the homozygous genotype for 24-bp mutation has significantly reduced chitinase activity [122–124]. These observations are in stark contrast with the in vitro data showing that recombinant CHIT1 can digest the fungal cell wall and lead to the death of fungus [100, 125]. However, we should be aware that the true roles of chitinase may still be unknown. Similarly, it is possible that the role of chitinases can be observed only in the presence of certain stress or disease conditions, rather than at baseline. As described in the next section, there are probably more unknown than known functions in chitinase biology that merit further investigation.
Unknowns in chitinase biology
One prominent question that still remains unanswered is the precise role of CHIT1 in mammals including humans and whether these proteins have any major role in the host. It is not clear if the 24-bp mutation is indeed an evolutionary beginning of loss of this gene function due to its relative uselessness. The cost of producing the protein at such high levels must be justified with a significant survival advantage in the face of a major threat faced by humans. It is currently not clear if CHIT1 function depends on its enzymatic activity. This is an important aspect of chitinase biology, as a significant population with 24-bp mutation has no chitinase activity but does have the molecule. A precise receptor for CHIT1 remains unknown, but has been identified for the related chitinase-3 like protein 1 which binds to IL-13Ra2 and requires TMEM219 [125–128]. Another major lack of knowledge regarding chitinase biology is our lack of understanding of the internal substrate for the chitinase enzyme in mammals. This is another major direction for potential further investigations. In the absence of a known receptor, our knowledge of specific pathways remains unknown.
In summary, mammalian chitinase is an enzyme with chitinolytic activity. The upregulation of the CHIT1 gene results in increased chitinase activity in many lung diseases; however, conflicting reports often emerge. Increased CHIT1 activity has been shown to play important roles in chronic pulmonary inflammation, allergy, injury, healing and pathogen infection. The effects of CHIT1 appear to be influenced by many factors, and its distribution and expression very between diseases. Much more research is needed to understand the precise roles of CHIT1 in biological functions or to characterise it as an important biomarker for lung disease severity.
Acknowledgement
We acknowledge the contribution of Liang Liu (3rd Medical Center of Chinese PLA General Hospital, Beijing, China) for the critical reviews.
Footnotes
Submitted article, peer reviewed.
Conflict of interest: D. Chang has nothing to disclose.
Conflict of interest: L. Sharma has nothing to disclose.
Conflict of interest: C.S. Dela Cruz has nothing to disclose.
Support statement: The authors gratefully acknowledge financial support from the China Scholarship Council (DC; 201809112037), Beijing Nova Program (DC; Z171100001117012 and 191100001119021), National Institutes of Health National Heart, Lung, and Blood Institute (CDC; R01HL126094), American Legion Auxiliary (LS; 513385) and the American Thoracic Society (LS: 2018-18). Funding information for this article has been deposited with the Crossref Funder Registry.
- Received October 29, 2019.
- Accepted December 2, 2019.
- Copyright ©ERS 2020.
This article is open access and distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0.











{kind=link}
{kind=link}
{kind=link}