





Abstract
Influx of eosinophils in airway mucosa and airway lumen is a hallmark of bronchial asthma. In-vitro data and animal studies indicate that the T‐helper (Th) type‐2 cell cytokine, interleukin (IL)‐5, plays an important role in eosinophil maturation, differentiation, recruitment, and survival. The objective of this study was to determine whether intravenous treatment with monoclonal anti-IL‐5 would affect the number of peripheral blood eosinophils, their activation status, T‐cell activation or the pattern of Th1 and Th2 cytokine production.
Over a period of 6 months, 19 asthmatics were investigated in a double-blind, placebo-controlled, multicentre study with mepolizumab (SB 240563) anti-IL‐5 antibody administered three times. Before each infusion and 12 weeks after the last infusion, peripheral blood leukocytes were examined, qualitative and quantitative distribution of eosinophils and lymphocyte subpopulations, frequencies of IL‐2, ‐3, ‐4, ‐5, -10, -13, interferon‐γ‐producing CD4 T‐cells and serum eosinophil cationic protein (ECP) levels were determined.
Treatment with mepolizumab resulted in a marked, rapid and sustained decrease of eosinophil numbers (median values from 300 to 45 per µL) paralleled by decreased levels of serum ECP (median values from 15 to 5 µg·L−1). Distribution of T‐cell subsets and T‐cell cytokine production were not altered during antibody treatment.
In conclusion, administration of mepolizumab to asthmatic patients markedly reduces peripheral blood eosinophils without altering the distribution and activation status of lymphocytes.
This study was supported in part by a grant from SmithKline Beecham, Harlow, UK.
Infiltration of the airways by CD4+ T‐helper (Th) type‐2 cells and eosinophils represents a predominant feature of the late-phase asthmatic response. The presence of these cells and their inflammatory products in the lung often correlates with disease severity and the degree of airway hyperreactivity 1, 2. The Th2 cytokine interleukin (IL)‐5 has been implied to play a pivotal role in inflammatory responses in animal models of allergic asthma 3, 4. IL‐5 plays an important role in development, recruitment, activation and survival of eosinophils 5–8. An obligatory role of IL‐5 was demonstrated in IL‐5‐deficient mice (IL‐5‐/‐) for the induction of eosinophilia and for the development of airway hyperreactivity to β‐methacholine in response to aeroallergen challenge 7. IL‐5 and associated eosinophilia regulate the induction of airway hyperreactivity during allergic inflammation in IL‐4‐deficient mice (IL‐4‐/‐) 9. Allergen inhalation increases local production of IL‐5 in the airways 10 as well as the number of peripheral blood eosinophils and lymphocytes (CD4+, CD8+ and CD4-/CD8- cells) containing intracellular IL‐5 11. In asthmatic patients, IL‐5 could be involved in eosinophilic inflammation since there are raised concentrations of IL‐5 in serum and increased numbers of IL‐5 messenger ribonucleic acid positive cells in bronchial biopsy samples 12, 13. In addition, raised levels of serum IL‐5 and blood eosinophils are associated with the fall in forced expiratory volume in one second (FEV1) that constitutes the late asthmatic reaction 14. Direct evidence that IL‐5 may play a main role in asthma originates from inhalation of IL‐5 by asthmatic patients, which causes hyperresponsiveness and sputum eosinophilia 15.
Therefore, the immunomodulatory functions of IL‐5 have been identified as a key therapeutic target. A murine-neutralising antibody to IL‐5 inhibits eosinophil infiltration into airways and lung tissue following allergen challenges in sensitised mice, guinea-pigs, and monkeys 16–18. In a guinea-pig model, hyperresponsiveness is blocked by treatment with anti-IL‐5 antibody prior to the challenge 17. A humanised anti-IL‐5 antibody (Sch 55700) has been constructed inhibiting pulmonary eosinophilia in guinea-pigs and primates 19.
The development of humanised monoclonal anti-IL‐5 antibodies provides the unique opportunity to investigate the contribution of IL‐5 to the pathogenesis of bronchial asthma in humans under in-vivo conditions. In a recent study, using the same monoclonal antibody, it was shown that a single intravenous application of anti-IL‐5 antibody reduced circulating eosinophils and sputum eosinophilia after allergen challenge 20. The current study aimed to assess immunobiological effects of the anti-IL‐5 antibody following repeated treatments. The effects on the biology of eosinophils, the levels of eosinophilic cationic protein (ECP) and basophils, in addition to the distribution, activation status and cytokine production of T‐cells were examined. Studies performed in murine models of experimental asthma revealed that genetic or pharmacological removal of IL‐5 prevents airway inflammation characterised by an influx of Th2 cells 7, 16. Although T‐cells lack significant expression of IL‐5 receptors, this observation raised the question that blockade of IL‐5 could have an impact on the Th1/Th2 profile in these patients over a longer period of time. Therefore, whether treatment with anti-IL‐5 would have any impact on the level of Th2 cells in peripheral blood was examined. This study was performed in a subgroup of patients participating in a multicentre study to investigate clinical effects of treatment with mepolizumab, a monoclonal anti-IL‐5 (SB 240563), in bronchial asthma.
Materials and methods
Study subjects
Seven male and 12 female patients with mild or moderate asthma, aged 20–59 yrs (mean 41 yrs), with duration of disease between 1–32 yrs (mean 11 yrs) were investigated. For inclusion, FEV1 had to be 50–80% of predicted at baseline, with a reversibility of ≥12%. None of the patients suffered from clinical exacerbation and all patients were on a stable daily dose of up to 1,000 µg beclomethasone diproprionate or a corresponding dose of other inhaled corticosteroids for ≥6 weeks prior to the study. As a symptom reliever salbutamol was allowed if needed. The detailed clinical characterisation of patients revealed no significant difference between the study groups (table 1⇓).
Patient characteristics
Study design
The clinical trial was performed over 6 months as a double-blind, randomised, placebo-controlled, parallel-group study with anti-IL‐5 antibodies (SB 240563, 250 mg or 750 mg per dose) administered intravenously. A run-in period of 1 month preceded three intravenous drug applications every 4 weeks, followed by a follow-up period of 3 months (fig. 1⇓). The study was performed according to the guidelines on good clinical practice and was approved by the local ethics committee. All participants gave their informed consent to this study. Ethylenediamine tetraacetic acid (EDTA) blood samples were drawn from the cubital vein ∼1 h before the first, 1 week after the second, 1 week after the third, and 3 months after the last infusion (fig. 1⇓) of SB 240563, for determination of absolute and differential cell counts, ECP levels and for flow cytometric analysis of lymphocyte subsets.

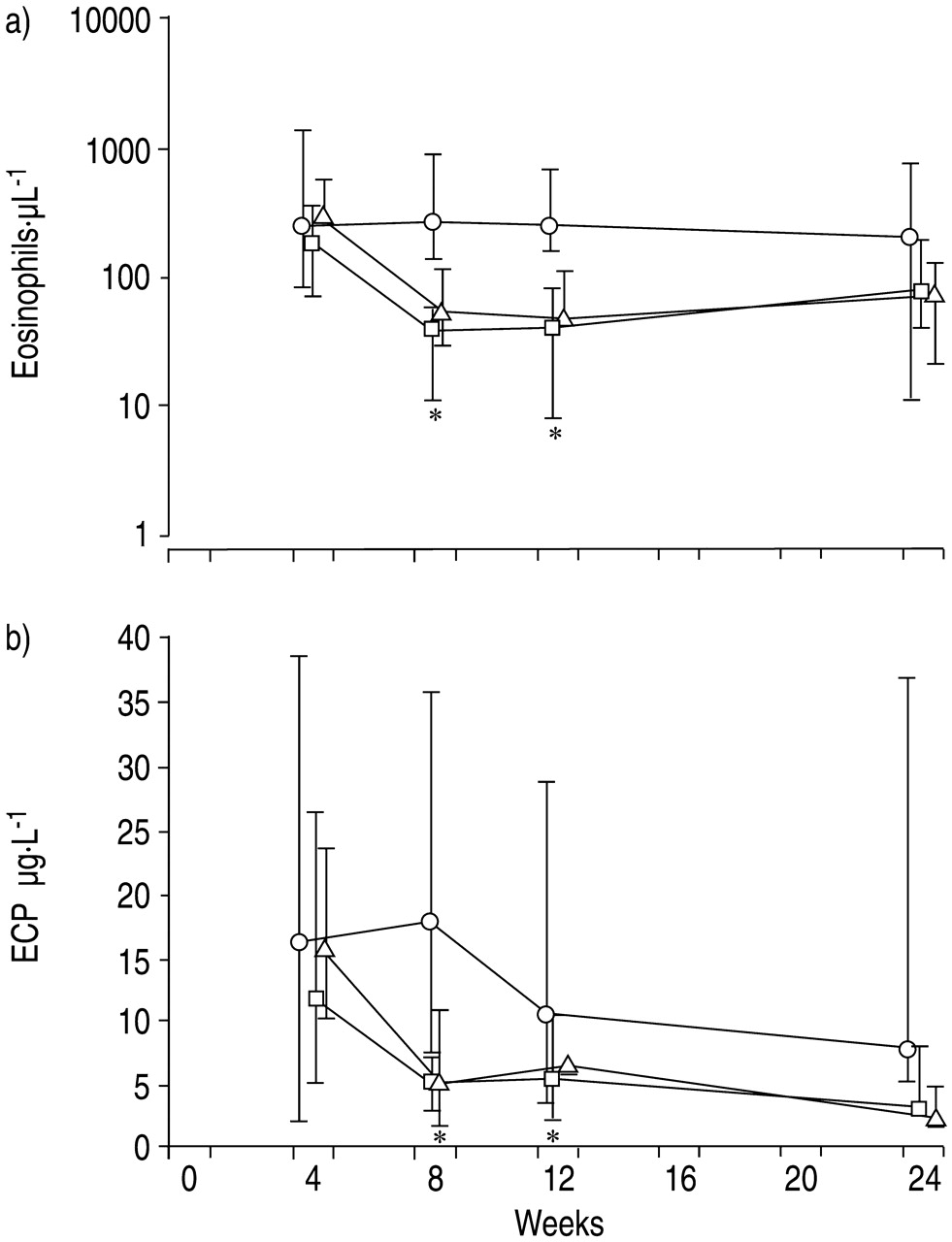
Time course of a) eosinophil granulocytes and b) eosinophilic cationic protein (ECP) in asthmatics treated with monoclonal antibody to interleukin‐5 at 4, 8 and 12 weeks. ○: placebo; □: 250 mg mepolizumab; ▵: 750 mg mepolizumab. *: p<0.05 between placebo and mepolizumab groups.
Methods
All samples were processed within 2 h of collection. Leukocyte count and their differentiation was determined routinely by the haematology analyser H3 (Bayer Diagnostics, Terrytown, NY, USA). Differentiation of lymphocyte subpopulations, expression of activation markers and intracellular cytokine production, as well as eosinophil activation, were assessed by flow cytometry using the fluorescence-activated cell sorter, FACScan (Becton Dickinson/Pharmingen, San Diego, CA, USA). Acquisition and analysis of data was performed as described previously 21. The lymphocyte gate was set, based on forward angle and 90° side-scatter, and purity was assessed based on CD45+/CD14- cells within the gate 21. The following markers were determined with phycoerythrin or fluorescein isothiocyanate-labelled antibodies (Becton Dickinson/Pharmingen) to CD3 or CD4 or CD8: CD19, CD16/56, CD45RA (naive) and RO (memory), γδ‐T‐cells, and cell activation by expression of human leukocyte antigen (HLA)-DR and CD25. SimulsetTM Controlγ1/γ2 (Becton Dickinson/Pharmingen) was used as negative control.
Flow cytometric assessment of T‐cell cytokine production was based on the stimulation of T‐cells by phorbolester plus ionomycin in the presence of monensin. Cells were stimulated for 6 h followed by fixation with paraformaldehyde, permeabilisation with saponin, and intracytoplasmatic staining of the accumulated cytokines IL‐2, ‐4, ‐5, -10, -13, or interferon (IFN)‐γ as described previously 22. Additionally, the expression of the IL‐4 and IL‐5 receptors, CD124 and CD125, respectively, and markers of cell activation (CD69, CD11b) on eosinophils were analysed. Further, the fraction of apoptotic eosinophil cells was determined by propidium iodide. This fraction was always <5% in all the samples.
Serum levels of ECP were determined by capacity radioallergosorbent test according to the guidelines of the manufacturer (Pharmacia, Freiburg, Germany). EDTA blood was centrifuged and plasma aliquots were frozen at −80°C within 2 h of blood collection. All samples were analysed in one assay.
Analysis
The significance of differences between the study groups was tested using the parameter-free Mann-Whitney test and a significance level of p<0.05 was used. In addition to absolute values at different time points, the time course was estimated by calculating individual differences between the values prior to and after drug application. Statistical data was represented as median and minimal and maximal values since the values were not distributed normally.
Results
Asthmatic patients received three consecutive intravenous infusions of either mepolizumab (250 mg or 750 mg per dose) or placebo at 4‐week intervals (fig. 1⇑). Remarkably, almost a complete disappearance of peripheral blood eosinophils was observed after the first infusion (fig. 1a⇑). Eosinophil counts remained low or absent until week 24, 12 weeks after the last infusion. In contrast, there were no significant changes in eosinophil counts in the placebo group. The marked fall in peripheral blood eosinophils was accompanied by a significant decrease in ECP concentrations (fig. 1b⇑). The kinetics of ECP levels resembled the eosinophil counts. These qualitative and quantitative changes were observed in both treatment groups without a significant difference between the 250 and 750 mg dosage.
Other markers of eosinophil activation (CD11b, CD69) or Th2 cytokine-receptor expression (CD125, CD124) remained unchanged (fig. 2⇓). Similarly to eosinophils, the basophil counts exhibited a moderate decrease in the 250 mg mepolizumab group (fig. 2⇓). Th cells, their activation markers (CD25, HLA-DR), and the Th2/Th1 intracellular cytokine pattern (IL‐4, ‐5, -10, -13/IFN‐γ, IL‐2) remained uninfluenced by IL‐5 suppression in treatment versus placebo groups (fig. 2⇓).

{kind=link}
{kind=link}
Time course of markers of eosinophil activation, T‐helper type‐2 cell cytokine-receptor expression, and peripheral blood lymphocytes in asthmatics treated with monoclonal antibody to interleukin (IL)‐5 at 4, 8 and 12 weeks. a) CD11, b) CD69, c) CD125, and d) CD124 on eosinophils, and e) basophil granulocytes, f) CD4 T‐cells, g) CD25 on CD4 cells, h) human leukocyte antigen-DR on CD4 cells, i) IL‐4‐positive CD4 cells, j) IL‐5‐positive CD4 cells, k) IL-10-positive CD4 cells, l) IL-13-positive CD4 cells, m) interferon‐γ‐positive CD4 cells and n) IL‐2‐positive CD4 cells. ○: placebo; □, ▵: 250 mg, 750 mg mepolizumab. #: p<0.05 between placebo and verum 250 of individual time course (D‐values pre- to noted time point).
The authors also examined whether anti-IL‐5 treatment would have any effect on other peripheral blood lymphocytes. The analyses included assessment of B‐cells, natural killer cells, CD3 and CD8 T‐cells, memory cells, naive cells, and γδ‐T‐cells. None of these parameters revealed a significant difference between study groups during the time of investigation (data not shown).
Discussion
This study clarifies the role of IL‐5 in the immunobiology of eosinophils in asthmatic patients. Blockage of IL‐5 by the corresponding antibody not only dramatically reduced the number of eosinophils, but also suppressed eosinophil activation as reflected by a marked reduction in ECP levels. This effect occurred without affecting T‐cell activation, distribution of their subpopulations, or their pattern of cytokine production. Thus, feedback mechanisms of anti-IL‐5 treatment on Th2/Th1 regulation seem to be of minor importance.
Numerous experimental data indicate the importance of IL‐5 for differentiation, maturation, growth, and survival of eosinophils 5–8. For example, animal models demonstrated the pivotal role of IL‐5 for terminal differentiation and mobilisation of eosinophils from the bone marrow into the circulation 8. The present authors' findings are in line with recently published data 20, which also showed a marked suppression of circulating eosinophils and a reduction of sputum eosinophilia after allergen challenge following a single intravenous administration of mepolizumab, the same monoclonal anti-IL‐5‐antibody used in this study 20. In a similar manner to Leckie et al. 20, the marked reduction of eosinophils occurred rapidly after the first antibody administration. In the present study, the authors were able to extend this observation by demonstrating that this effect persisted for at least 12 weeks after termination of antibody treatment. Furthermore, they observed a strong reduction of serum ECP levels in parallel to the decline in eosinophil numbers.
Reasons for the disappearance of eosinophils from peripheral blood may be either the suppression in eosinophil maturation and differentiation in the bone marrow or an increased influx of eosinophils into the inflamed tissue. Based on the previous observation of a reduction of sputum eosinophilia after allergen challenges following antibody treatment 20, it is more likely that the antibody indeed inhibited maturation and differentiation of the cells in the bone marrow. An increased influx of eosinophil into tissues, however, cannot be formally ruled out by these data.
Even after repeated antibody infusions, neither the frequencies of “naïve” (CD45RA) and “memory” (CD45RO) T‐cells nor the frequencies of cytokine-producing CD4 T‐cells were altered in any way. These data strongly support the concept that although IL‐5 is an important cytokine produced by Th2 T‐cells, it does not affect the activation status of mature T‐cells and does not alter the pattern of Th1 (as reflected by the frequencies of IL‐2‐ and IFN‐γ‐producing CD4 T‐cells) and Th2 (IL‐3‐, ‐4‐, ‐5‐, -10- and -13-producing CD4 T‐cells) cytokine production. These data are in line with the observation that T‐cells do not express significant levels of IL‐5 receptors. Furthermore, even if anti-IL‐5 treatment would have a local anti-inflammatory effect, suggested by experimental murine models of asthma, this would not affect the Th1/Th2 profile in these patients.
Over the last two decades, the concept has been developed that CD4 T‐cells and eosinophils play an important part in the (immuno)pathogenesis of bronchial asthma 1–3, 23, 24. An influx of both CD4 T‐cells and eosinophils has been observed in bronchial tissue, as well as into the airway lumen, particularly in late-phase allergic responses 25. However, the contribution of these cells to the development of bronchial hyperresponsiveness and the decline in lung functions is still a matter of intense investigation 3, 9, 10, 26, 27. Even in mice, the contribution of IL‐5 on airway hyperresponsiveness has been controversial 7, 16, 26. There is increasing evidence for the interaction between airway eosinophils and CD4 T‐cells via cytokines other than IL‐5, i.e. IL-13, which may be of great importance for the development of asthmatic symptoms and activation of eosinophils 28, 29. Furthermore, the contribution of other immune cells in addition to eosinophils and T‐cells may have been underestimated.
The present study investigated the effect of single intravenous application of mepolizumab and there was no measurable effect on allergen-induced, late-phase responses or on the degree of airway hyperresponsiveness. The findings lead the authors to conclude that eosinophils may not have the relevance to the pathogenesis and treatment of asthma as assumed previously. In this regard, it needs to be pointed out that this study was not designed to correlate anti-IL‐5 treatment with clinical outcome parameters. These data will, however, be provided in the context of the entire multicentre study currently under assessment.
These experimental data clarify the role of interleukin‐5 in the in-vivo immunobiology of eosinophils in asthmatic patients. The exact role of interleukin‐5 and eosinophils in the pathogenesis of bronchial asthma, particularly with respect to clinical hallmarks, such as airway hyperresponsiveness, reversible broncho-obstruction and airway remodelling, needs to be investigated.
Acknowledgments
The authors would like to thank U. Meysel, C. Dumont, and E. Hungerbühler for their excellent technical assistance.
- Received April 3, 2002.
- Accepted January 24, 2003.
- © ERS Journals Ltd









